The following is a major assignment of mine (in two parts because of its length) on the Cambrian Explosion, which was part of an animal physiology unit in the last semester of my biology degree. I had a choice in which assignment, but not the heading. That is, we had about 30 assignment topics to chose from, and my policy always was that, to maximise the benefit of my time at university, where I had a choice, to chose the most `evolutionary' topic I could.
I am posting this, not because it is a publication-quality paper (it isn't-although I did get a high mark for it), but because it represents (or did in 2004) the latest main lines of evidence (within the space and time constraints of a mere undergraduate assignment), on the Cambrian Explosion. As can be seen in part #2, it has over 70 references, most being scientific journals. While there is nothing I wrote here that I don't believe is true (except the mandatory calling it "evolution"-though I tried to do that as least as possible, not just here but all though the degree), I obviously could not say (without being failed by my atheist lecturer) that God created, i.e. in the sense of "mediate, progressive creation" (which is what I believe):
"Mediate and Immediate Creation. But while it has ever been the doctrine of the Church that God created the universe out of nothing by the word of his power, which creation was instantaneous and immediate, i.e., without the intervention of any second causes; yet it has generally been admitted that this is to be understood only of the original call of matter into existence. Theologians have, therefore, distinguished between a first and second, or immediate and mediate creation. The one was instantaneous, the other gradual; the one precludes the idea of any preexisting substance, and of cooperation, the other admits and implies both. There is evident ground for this distinction in the Mosaic account of the creation. ... It thus appears that forming out of preexisting material comes within the Scriptural idea of creating. ... There is, therefore, according to the Scriptures, not only an immediate, instantaneous creation ex nihilo by the simple word of God, but a mediate, progressive creation; the power of God working in union with second causes." (Hodge C., "Systematic Theology," [1892], James Clark & Co: London, Vol. I, 1960, reprint, pp.556-557. Emphasis original)
But with my Christian creationist eyes I saw abundant of evidence of the providential working on a grand scale of Him who has a "plan determined for the whole world" (Isaiah 14:26); "who works out everything in conformity with the purpose of his will" (Ephesians 1:11). However, as can be seen, I did have a shot at Darwin. And as this assignment was at first a slide presentation to my fellow students, it was probably the only criticism of Darwin they had ever heard in their biology degree!
Stephen E. Jones, 1 November 2004
INTRODUCTION
The Cambrian evolution of animal body plans
As the title of this paper suggests, the evolution of animal body plans took place in the Cambrian period (Fig.1), which was between 543 and 490 Ma (Conway Morris, 2002; Knoll, Walter, Narbonne & Christie-Blick, 2004). In fact, more precise radiometric dating has 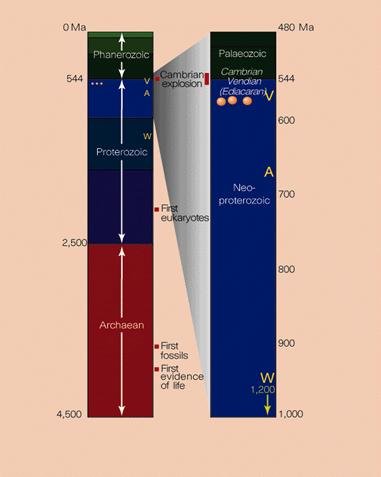 revealed that this evolution of animal body plans took place between 533 and 525 Ma (Bowring, et al., 1993; Kerr, 1993). This was a maximum of 8 million years, but more likely it was between only 5 to 6 million years (Carroll, 1997, p.4; Gould, 1994b), which is less than 1% of life’s ~3,800 Ma history (Mojzsis, et al., 1996).
revealed that this evolution of animal body plans took place between 533 and 525 Ma (Bowring, et al., 1993; Kerr, 1993). This was a maximum of 8 million years, but more likely it was between only 5 to 6 million years (Carroll, 1997, p.4; Gould, 1994b), which is less than 1% of life’s ~3,800 Ma history (Mojzsis, et al., 1996).
Fig. 1. Geological timescale of the Cambrian explosion. Source: Bengtson, 1998.
Cambrian explosion
So geologically sudden and explosive was this Cambrian evolution of animal body plans, that it is called "the Cambrian explosion" (Fig. 1), or evolution’s "big bang" (Conway Morris, 2002; Kerr 1993; Levinton, 1992). The Cambrian explosion is rightly regarded as one of the most dramatic and important event in the history of animal life (Bowring, et al., 1993; Hickman, Roberts & Larson, 2000, p.36; Orr, Briggs & Kearns, 1998).
Aim
The aim of this paper is to examine the evidence for the evolution of animal body plans in the Cambrian and immediately preceding Ediacaran periods, and the main proposed causal explanations. The title elements will be used as a framework, but in a different order.
ANIMAL BODY PLANS
Metazoan triploblasts
By "animal" is meant the triploblastic metazoa. The metazoa are complex multicellular animals, with cellular, tissue and organ levels of organisation (Conway Morris, 1998, p.24; Hickman, Roberts & Larson, 2000, p.38). In the five-kingdom classification system of life, Metazoa are equivalent to the kingdom Animalia (Margulis, Schwartz & Dolan, 1994, p.14). The Metazoa include both diploblastic and triploblastic animals with two and three primary layers of body tissue, respectively (Gould, 1998). Modern animals, except for sponges, corals, and some minor groups, are triploblastic, with three body tissue layers, enabling the development of complex, mobile, bilaterally symmetrical animals with body cavities, gut, anterior-posterior orientation, paired appendages and sensory organs (Gould, 1998). The Cambrian evolution of animal body plans here refers only to triploblastic animals (Conway Morris, 2002).
Phyla
By "body plans" is meant the fundamental anatomic ground- plan, which is the basis of each phylum; the basic unit of classification within each kingdom (Gould, 1989, p.99). There are generally regarded as being between 32 and 35 animal phyla existing today, with each phylum characterized by a distinctive body plan that sets it apart from all other phyla (Hickman, Roberts & Larson, 2000, p.36; Raff, 1999). Unless otherwise indicated, from now by "animal" and "animal phyla" is here meant triploblastic animals.
In the Cambrian explosion, all of the animal phyla, together with some novel body plans that we know only from the fossil record, were established (Carroll, 1997, p.344; Hickman, Roberts & Larson, 2000, p.36). While the bryozoan phylum first appeared in the next Ordovician period, and a few new phyla have been discovered since, it is generally assumed because of their small size and on phylogenetic considerations, that these reflect a failure to find (Gould, 1994b; Giribet, 2002). The Cambrian fossil assemblages are dominated by invertebrate phyla such as annelids, molluscs, arthropods, brachiopods, and echinoderms (Conway Morris, 1998, p.30; Shu, et al., 1999). However, chordates are also represented (Chen, Huang & Li, 1999; Chen, et al., 2003).
Just as surprising as the sudden appearance of the animal phyla, is the lack of new animal body plans to emerge since that time (Gould, 1994b; Holmes, 1997). All the evolutionary changes in animals since the Cambrian, even the bursts of speciation that followed major extinctions, produced only variations on those established themes (Gould, 1994a; Levinton, 1992). For example, about 80% of all described animal species are arthropods, and of those, most are insects (Gould, 1989, p.49). The main reason for this extreme conservatism seems to be that once established, a basic body plan becomes an architectural constraint for all descendants of that ancestral line (Hickman, Roberts & Larson, 2000, p.36; 1992).
CAMBRIAN
Darwin’s "difficulty" (1859)
That Precambrian strata did not contain animal fossils was evident to geologists by the 1830s (Conway Morris, 1998, p.140). This apparent sudden beginning of complex animal life in the geological strata of the Cambrian Period was a major difficulty for Darwin, who in his 1859 Origin of Species wrote of it as a
"difficulty, which is much more serious. ... the manner in which species belonging to several of the main divisions of the animal kingdom suddenly appear in the lowest known fossiliferous rocks. if the theory be true, it is indisputable that before the lowest Cambrian stratum was deposited ... the world swarmed with living creatures. ... why we do not find rich fossiliferous deposits belonging to these assumed earliest periods prior to the Cambrian system, I can give no satisfactory answer" (Darwin, 1872, pp.314-315).
The problem for Darwin’s theory of gradualistic evolution was that in his day there were thought to be Precambrian fossils of a simple organism named Eozoon (which later turned out to be an inorganic mineral deposit), and also it was known that there were comparatively advanced trilobite, mollusc and annelid fossils in the Cambrian strata, but no transitional fossils in between (Darwin, 1872, pp.316, 447; Gould, 1980, pp.198-201).
However, since Darwin’s day, fossil and other evidence has been discovered that, while it has vindicated his expectation of the Precambrian world swarming with animal life, it has not matched his theory’s prediction of a gradualistic increase in the complexity of animal life through the Precambrian towards the Cambrian (Carroll, 1997, p.2; Gould, 1989, p.57).
Evidence of Cambrian life
Burgess Shale (~505 Ma, Canada)
The Burgess Shale (Fig. 2) in Canada was discovered in 1909 by Charles Walcott of the Smithsonian Institution, and was the first known major assemblage of Cambrian fossil fauna (Conway Morris, 1998, p.18; Gould, 1989, p.23). At ~530 Ma the Burgess Shale represents the period just after the Cambrian explosion, and so preserved a wide range of its fossils in exquisite detail,  including impressions of their soft anatomy (Gould, 1989, p.23).
including impressions of their soft anatomy (Gould, 1989, p.23).
Fig. 2. Burgess Shale fauna. Anomalocaris has captured a trilobite. Source: The William Younger Centre, 2000; Conway Morris, 1998, p.88a.
Chengjiang (~515 Ma China)
The Chengjiang biota from China is a diverse assemblage of Early Cambrian marine fossils, comparable to, but earlier than, the Burgess Shale and preserving in the same exquisite detail not only the organisms’ hard skeletal parts but also their soft parts (Babcock, 2001). Other Cambrian fossil beds have been discovered, for example Sirius Passet in Greenland, which is about the same age as Chengjiang (Conway Morris, 1998, p.xxi).
Cambrian phosphatized fossils
In 1985, Upper Cambrian tiny crustacean larvae, preserved by a process called phosphatization, were reported found in Sweden (Gould, 1998). Phosphatization, the replacement of tissue with calcium phosphate, was a product of the unusual geochemistry of the Proterozoic-Cambrian transitional period, but it works only for tiny objects up to about 2 mm (Budd, 2004). Preservation by phosphatization is often in exquisite, three-dimensional detail, revealing soft anatomy, bristles, hairs, and even cellular structures (Bengtson, 1998; Gould, 1998; Kerr, 1998a). The discovery in Middle Cambrian (~510 Ma) strata of phosphatized embryonic cells representing the earliest stages of trilobites was reported in 1994 (Zhang & Pratt, 1994). In 1997, phosphatized embryos from the Early Cambrian were reported to have been traced in a growth series-from embryos to tiny near adults for an extinct group and a segmented worm (Bengtson & Zhao, 1997). A tiny 330-340 µm long phosphatized ostracod crustacean with appendages was in 2001 reported from the Lower Cambrian of England (Siveter, Williams & Waloszek, 2001). That such a relatively advanced arthropod as a crustacean appeared in the Early Cambrian means either it evolved with exception rapidity, or it supports the hypothesis of a long Precambrian "fuse" for the Cambrian  explosion (Fortey, 2001). A series of phosphatized embryos preserving developmental stages from cleavage to pre-hatching larvae (Fig 3) was reported in 2004 from Middle-Late Cambrian strata in Hunan, China (Dong, Donoghue, Cheng & Liu, 2004).
explosion (Fortey, 2001). A series of phosphatized embryos preserving developmental stages from cleavage to pre-hatching larvae (Fig 3) was reported in 2004 from Middle-Late Cambrian strata in Hunan, China (Dong, Donoghue, Cheng & Liu, 2004).
Fig. 3 Phosphatized embryo of Cambrian worm Markuelia hunanensis. Source: Dong, et al., 2004.
Evidence of Precambrian life
Ediacaran biota (~545 Ma, Australia)
The Ediacaran biota (Fig. 4), representing the first known major Precambrian fossil assemblage, was discovered by an Australian geologist R.C. Spring in 1946, in the Ediacara hills north of South Australia (Levinton, 1992, p.54; Sprigg, 1947). Ediacaran fossil assemblages have since been found world-wide, including Namibia, the White Sea coast of Russia, Newfoundland, China, Mexico and Canada (Conway Morris, 1998, p.25). While the classification of many Ediacaran fossils is still controversial, with some claiming they were "failed experiments" and others that they were frond-like symbionts with photosynthetic cyanobacteria, the Ediacaran biota does contain the oldest animals (Daviss, 1998; Knoll & Carroll, 1999; Raff, 1999). However, these Ediacaran animals are mostly diploblastic, whereas the Cambrian explosion was of triploblastic animals (Gould, 1998). The Ediacarans are therefore not a solution to Darwin’s difficulty, because they are only barely Precambrian, their relationship to the Cambrian fauna is problematic and they are soft-bodied with a worldwide distribution, therefore it cannot be claimed  that the ancestors of the Cambrian animals simply lacked hard parts and so did not fossilise (Li, Chen & Hua, 1998; Gould, 1989, p.59).
that the ancestors of the Cambrian animals simply lacked hard parts and so did not fossilise (Li, Chen & Hua, 1998; Gould, 1989, p.59).
Fig. 4 Ediacaran fossil Dickinsonia, an annelid. Source: University of California Museum of Paleontology.
Microfossils (~3.4 Ga - Australia)
In 1993 William Schopf discovered fossilized microorganisms in rocks 3.4 Ga in the Pilbara region of Western Australia (Schopf, 1993). However, it has been challenged that these are not microfossils, but only inorganic chemicals deposited by a hydrothermal vent (Rai & Gautam, 1999; Kerr, 2002). But a recent study testing these hypotheses concluded that the deposition occurred in normal open shallow to deep marine environments, with no evidence that hydrothermal processes had any direct role and therefore photosynthetic organisms were indeed living 3.4 Ga (Tice & Lowe, 2004).
Trace fossil burrows (~1 Ga & ~550 Ma)
Evidence of burrowing trace fossils ~1 Ga has been claimed (Seilacher, Bose & Pflüger, 1998). But this has been challenged as an inorganic pseudo-fossil (Kerr, 1998b; Rai & Gautam, 1999). However, undisputed trails and burrows have been found extending back to 550 Ma (Erwin & Davidson, 2002; Knoll & Carroll, 1999). Fossilised faecal pellets have also been found ~600 Ma (Knight, 1997). These trace fossils were evidently made by bilaterally symmetrical, worm-like animals with a mouth-anus digestive tract (Carroll, 2000).
Molecular "clock" estimates (~1.2 Ga - ~600 Ma)
Estimates for the actual origin of the animal phyla, as opposed to its first appearance in the fossil record, based on molecular "clock" divergences of seven genes, yielded a time of ~1.2 Ga for the divergence between chordates and arthropods, annelids, and mollusks (Gould, 1998; Holmes, 1997; Wray, Levinton & Shapiro, 1996). However, this has been challenged by other molecular "clock" estimates (Fig. 5) using different assumptions and molecules, including mitochondria and ribosomal RNA., that yield divergence times of between ~900 and ~600 Ma (Ayala, Rzhetsky & Ayala, 1998; Benton & Ayala, 2003; Bromham, Rambaut, Fortey, Cooper & Penny, 1998; Erwin & Davidson, 2002; Hedges, Blair, Venturi & Shoe, 2004). These times are still well before the Cambrian, and therefore have a problem explaining trace fossils which commence just before the Cambrian (Conway Morris, 1998, p.25). Their widely diverging estimates for the same events indicate that molecular "clocks" are less reliable for such distant and unusual evolutionary events (Conway Morris, 2002; Knoll, 2003). 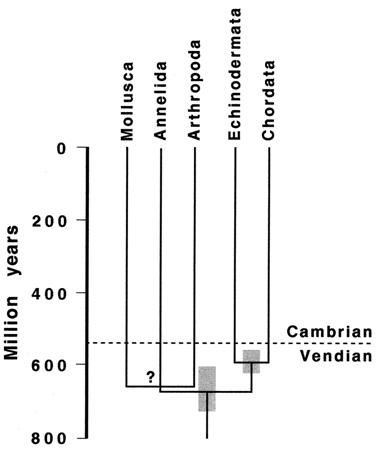 Nevertheless, all the molecular estimates agreed that metazoan divergence commenced well before the appearance of animal body plans in Cambrian explosion (Knoll, 2003). And they all seemed to agree that the original divergences of the phyla were almost simultaneous (Levinton, 1992).
Nevertheless, all the molecular estimates agreed that metazoan divergence commenced well before the appearance of animal body plans in Cambrian explosion (Knoll, 2003). And they all seemed to agree that the original divergences of the phyla were almost simultaneous (Levinton, 1992).
Fig. 5 Molecular "clock" phylogeny of protostome-deuterostome divergence ~670 Ma. Source: Ayala, Rzhetsky & Ayala, 1998.
Phosphatized Precambrian fossils (~560-600 Ma)
In 1998, Precambrian phosphatized sponges were reported as found 40-50 million years (~580 Ma) before the Cambrian explosion (Li, Chen & Hua, 1998). In the same year embryos of triploblastic animals from southern China were reported as found in ~570 Ma strata (Xiao, Zhang & Knoll, 1998). These embryos cannot be assigned to any particular group, but they strongly resemble embryos of living crustaceans (Gould, 1998; Knoll, 2003). Also, in 2000 phosphatized fossil embryos and larvae of cnidarians were reported as found in the Doushantuo Formation (~570 Ma) in China, although the claimed blastospheres are unusually large and some of the internal structures may be mineral growths (Chen, et al., 2000; Conway-Morris, 2003). Recently in 2004 it was reported that ten phosphatised fossils of small (<180 µm) but adult bilaterian animals dating between 560 - 600 Ma, or 40 - 55 Ma before the Cambrian explosion had been found in the Doushantuo Formation (Chen, et al., 2004). The animal (Vernanimalcula guizhouena) had a triploblastic structure, anterior-posterior organization, a gut within paired coeloms; possible paired sense organs, a pharynx and a ventrally directed anterior mouth. These fossils confirmed the phylogenetic inference that Bilateria arose well before the Cambrian  (Chen, et al., 2004).
(Chen, et al., 2004).
Fig. 6. Phosphatized Precambrian (~ 580-600 Ma) Bilaterian fossil, Vernanimalcula guizhouena. Source: Chen, et al., 2004).
Continued in part #2.
Stephen E. Jones, BSc (Biol).
"Problems of Evolution"



No comments:
Post a Comment